." In: Hanson G, Berliner LJ, eds.
. Vol. 12. Cham: Springer Publishers; 2017:.
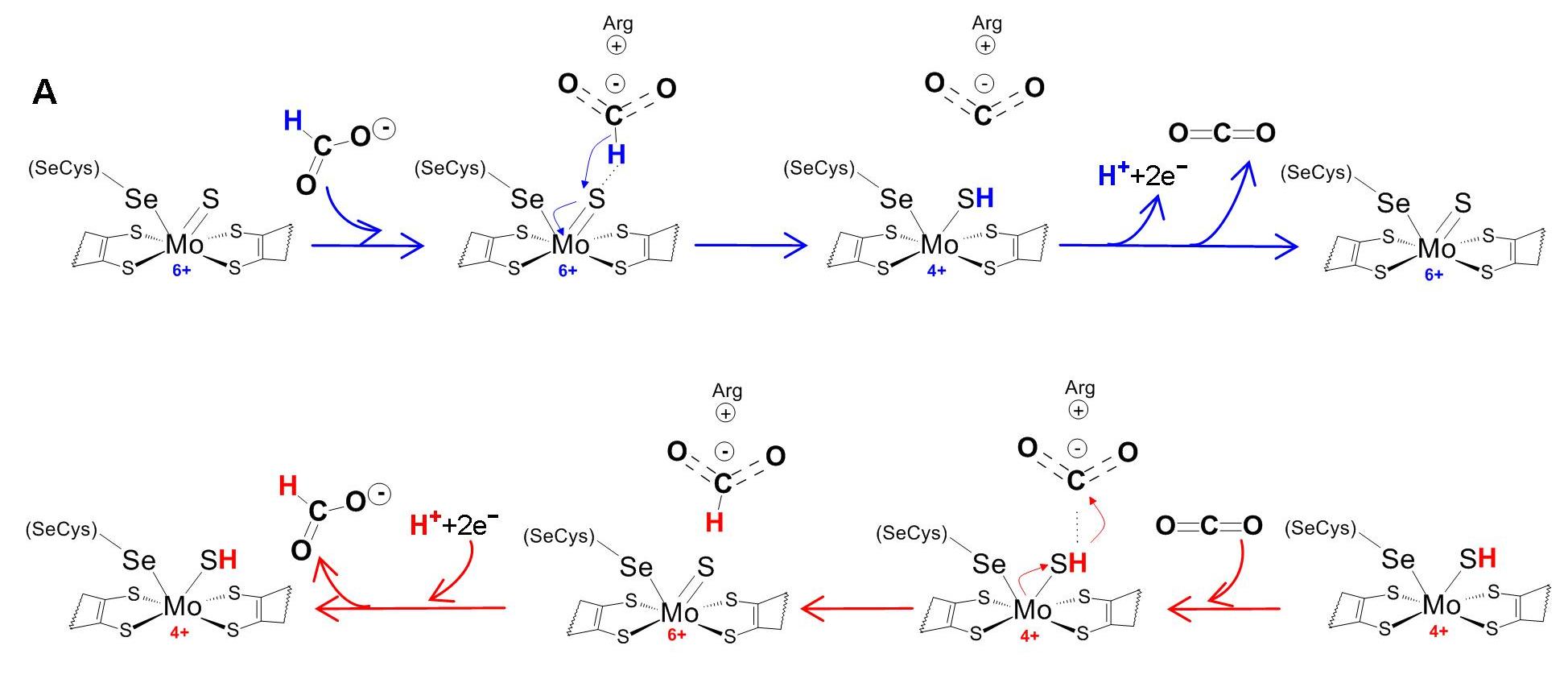
The biological relevance of molybdenum was demonstrated in the early 1950s-1960s, by Bray, Beinert, Lowe, Massey, Palmer, Ehrenberg, Pettersson, Vänngård, Hanson and others, with ground-breaking studies performed, precisely, by electron paramagnetic resonance (EPR) spectroscopy. Those earlier studies, aimed to investigate the mammalian xanthine oxidase and avian sulfite oxidase enzymes, demonstrated the surprising biological reduction of molybdenum to the paramagnetic Mo5+. Since then, EPR spectroscopy, alongside with other spectroscopic methods and X-ray crystallography, has contributed to our present detailed knowledge about the active site structures, catalytic mechanisms and structure/activity relationships of the molybdenum-containing enzymes.
This Chapter will provide a perspective on the contribution that EPR spectroscopy has made to some selected systems. After a brief overview on molybdoenzymes, the Chapter will be focused on the EPR studies of mammalian xanthine oxidase, with a brief account on the prokaryotic aldehyde oxidoreductase, nicotinate dehydrogenase and carbon monoxide dehydrogenase, vertebrate sulfite oxidase, and prokaryotic formate dehydrogenases and nitrate reductases.
http://docentes.fct.unl.pt/lblm/files/epr_book-chap_4.pdf
{kind=link}